Aging in Saccharomyces cerevisiae

Last updated 8/26/05
The current research of the Hoopes lab is concerned with the mechanisms of aging in the budding yeast. This research is supported by an NSF RUI and MRI grants and an NIH AREA grant, a Pomona College Research Committee grant to LH and also by various supporting stipends to her students, including Biology Department support, the Biology Department NSF REU site grant, the Pomona College SURP program, the Merck/AAAS Fellowship grant, and the Beckman Scholarship grant.
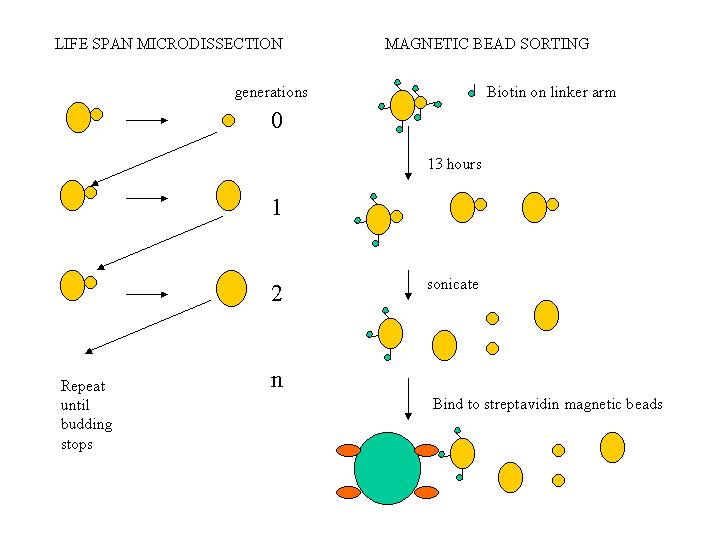
Yeast undergo a spiral form of cellular aging, where each mother cell appears at age zero generations and then undergoes budding for an average life span of 25-30 generations in a typical wild type strain. At each generation until the very end of life, the buds appear as zero generation cells; thus each cell division is assymetrical with respect to age. Mutations can increase or decrease the life span of a mother cell. The life span can be determined by microdissection, or cells of a particular number of generations can be isolated using biotinylation and streptavidin paramagnetic beads.
When we want to see what has happened to a cell near the end of its lifespan, we can look at its size, its replication rate, its nucleolar morphology (the sickle/cap-shaped nucleolus of young cells enlarges and fragments in older cells), the sterility or ability to respond to alpha factor to prepare for mating, and the amplification of rDNA. Our lab of wonderful undergraduates has collaborated in the past with that of Judith L. Campbell at CalTech in these studies. We have shown that dna2 mutants, which have alterations in an important (essential) DNA helicase for replication and repair, have short life spans. This is interesting since sgs1 deletions, taking out of commission a helicase homologous to that asssociated with Werner Syndrome (an early aging syndrome of humans), has been shown by Guarente's group to cause acceleration of aspects of yeast aging. We have looked at size increases, sterility, nucleolar fragmentation, and amplification of rDNA, and find all occur prematurely in the dna2 mutant strains. Since the DNA2 gene codes for a replication enzyme, involved in maturation of Okazaki fragments on the lagging strand, we tested the life spans of a number of replication mutants. In most cases, there was a statistically significant life span reduction; Jessica Brown '02 (MIT PhD program, NSF graduate fellowship) found that the pol1-14 mutant undergoes very early nucleolar fragmentation during aging. Our model for DNA aging is currently that there is replication stress resulting either from damage or replication fork stalling and that this stress, which occurs in normal aging, is intensified in replication mutants. Some of her work was used by LH to illustrate a talk at the ASM meeting in Salt Lake City in May, 2002 on use of genomics in undergradate research. Michelle Yuen '04 (University of Washington PhD program) looked at the role of Sir2p in the expression of genes in young and old cells, and in her experiments has found that some genes do change in expression early as a function of the dosage. She presented her work at the American Society for Microbiology in December, 2002 in San Francisco, and at the Salk Institute in August, 2002 at the DNA Replication and Genome Integrity meetings. Allen Kuo (MD program, Med College of New York) studied the early changes in gene expression during aging using microarrays, and has improved the magnetic sorting method to provide sufficient cells for the RNA needed in these experiments. His work on aging microarrays was continued by Michelle Yuen, Rishi Jindal ('04, Medical School), Jennifer Hardee ('05, Yale PhD program), and Laty Cahoon (UCLA '03, research technician, now at Northwestern PhD program).
WonChae Choe, who completed his dissertation at Caltech, showed that Dna2p is located at telomeres during much of the cell cycle in young cells. Kevin Pang ('00) at Pomona began to look at whether or not this association continues as cells age, and found indication that the protein began to move to the nucleolus rather early in the life span. This work was continued and extended to older ages by Selena Cataldo ('02), who presented a poster based on this work at the American Society for Microbiology meetings in Orlando, FL in May, 2001. She found that between 6 and 8 generations, a mother cell moved from a mixture of telomeric clump staining and nucleolar staining to virtually all nucleolar staining, and that this pattern was maintained up to 15 generations. While these cells are not really old (average life span is 25 generations), there are some cells that stop dividing as early as 13 generations, so it is reasonable to consider that they have undergone an aging process that is as yet incomplete.
Laura Hoopes has shown what appeared to be aging clock resetting in a very long-lived meiosis strain from Jim Haber's laboratory. The cells were aged and sorted before inducing meiosis; the life spans of aged vegatative, normal unaged vegatative, and postmeiotic aged cells were compared. The postmeiotic aged cells had life spans almost as long as the normal (unaged) cells; far longer than the aged cells had when grown on rich medium to prevent meiosis. But there was a deficit when compared with a normal, young life span. This experiment gave very similar results with the BR1919 strain from Shirleen Roeder and Beth Rockmill that is a good meiosis strain but has a shorter life span. In this case, Kyle Lancaster ('05, Caltech PhD program, NSF graduate fellowship)Meghan McFarlane ('02), Vinita Lukose ('08) and research associates Michelle Wu and Laty Cahoon found that the aging clock was reset meiosis of wild type and in ndt80 but not in spo11. Our collaborator, Beth Rockmill at Yale, has constructed strains for analysis with Zip1-GFP so we can easily detect when the cells reach pachytene stage of meiosis, where ndt80 stops the process.
In addition, the clock is reset in every budding cell division (mitosis). Sheila Vasantharam (law school) studied the role of asymmetry in this process, and has found that one gene that affects asymmetry also affects life span. Eventually, we plan to look for genes that regulate clock resetting in yeast, and hope to find homologues in higher organisms. Ranor Basu is considering studying this vegetative resetting for his senior thesis.
We expect that DNA replication problems will trigger checkpoint genes, and Alyssa van Dyk (MD program) studied the role of rad9 in the response of replication mutants to aging. Also, we found evidence earlier that rDNA amplification was occurring in aging cells, even if no large increases in ERCs were found. Kareen Wilkinson worked on analysis of chromosome XII via CHEF gel electrophoresis, and was not able to document any changes in chromosome size with age, though we have not been convinced that our technique would effectively detect such changes so far. Improvements in technique are being investigated. Additional studies on rDNA structure using 2D gel analysis were done by Michelle Yuen, Jennifer AhKee ('05, Fullbright to France and then PhD program at U Wisc, Madison), and Kristina Bajema ('06). Additional studies looking for DNA breakage via Comet assays, TUNEL assays, and QAOS PCR based analaysis have been done by Jennifer AhKee and Gloria Yiu ('08). The data to date support an increase in breakage by 8g.
Jennifer AhKee, Michelle Wu, and Laty Cahoon prepared samples for dynamic molecular combing to compare aging wild type and strains with altered doses of SIR2. Samples are sent to Philippe Pasero's laboratory in Montpellier for analysis by dynamic molecular combing. So far, data have indicated that origins of replication are farther apart in the old cells of wild type. Jennifer AhKee will carry on these experiments during her Fullbright fellowship in France.